1) instrument and interlock

仪表和连锁
2) instrument interlock protection
仪表连锁保护
3) Interlock Forms
连锁表
1.
Research on Automatic Generation of Interlock Forms for Train Route and Shunting Route Based on Graphic Structure;
基于图形结构的列车/调车进路连锁表的自动生成算法研究
4) interlocking account
连锁报表
5) locking list
连锁图表
6) instrument with locking device
锁定型仪表
补充资料:连锁和交换
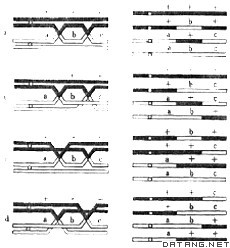
| 连锁和交换 linkage and crossingover 位于同一染色体上的基因伴同遗传的现象称为连锁。由于同源染色体相互之间发生交换而使原来在同一染色体上的基因不再伴同遗传的现象称为交换,在原核生物中多称为重组。连锁和交换是生物界的普遍现象。 连锁对于生命的延续是十分必要的。因为一个细胞中有许多基因,如果它们各个分散,便很难设想在细胞分裂过程中怎样可以使每一个子细胞都准确地获得每一个基因。交换对于生物的进化有重要意义,它可以使配子中的基因组合变化无穷,从而带来生物个体间的更多的变化,为自然选择提供更大的可能性。 交换一般是对等的,可是也有不对等的。不对等交换导致少量染色体重复,这被认为是生物进化中新基因的主要来源之一。此外,交换和育种工作也有密切的关系。重组机制的研究是遗传学中的一个古老的问题,现在已经成为分子遗传学研究中的吸引人的课题。 物质基础 1913年美国遗传学家A.H.斯特蒂文特首先在果蝇中测定了6个基因在X染色体上的排列顺序,获得了第一个连锁群,奠立了遗传的染色体学说。根据这个学说,可以预期连锁群数等于单倍染色体数。在深入研究过的生物中,都证实了这一假设。1909年F.A.詹森斯首先在两栖类动物和直翅目昆虫生殖母细胞的减数分裂中,看到了二价体中的交叉,并提出交叉型学说,认为交叉是交换的结果而不是交换发生的原因。也就是说遗传学上的交换发生在细胞学上的交叉出现以前,而并不是由于染色单体相重叠而导致交换的发生。按照交叉型学说,交换在粗线期甚或更早时期已经发生;在双线期中两个未分裂的着丝粒分别带着两个姊妹染色单体趋向两极时使已经发生交换的位置上出现交叉。 在光学显微镜下,交叉首先在双线期中看到,可是在电子显微镜下在偶线期-粗线期就可以看到称为联会线复合体的结构。它的显微图像在各种生物中不完全相同,但是基本上包括两条侧线和一条中轴,中轴和侧线之间往往可以看到许多横纹。许多事实都说明这一结构和联会与交叉有关。 发生的位置 一般情况下染色体愈长则显微镜下可以看到的交叉数愈多。但交叉数的多少又不完全决定于染色体的长度。在某些生物中,在某一长度范围内染色体长度和交叉数呈正比;如果低于这一长度范围,则实际交叉数比预期数大。例如根据染色体长度看来预期交叉数不到一个的二价体上都出现了一个交叉。这说明任何一个染色体不论它是怎样短,都必然发生一个交叉。这种机制现在还不清楚,一般认为它对于细胞分裂过程中染色体的有规律的行为起着重要的作用。 在某些生物中,交叉在染色体上的分布倾向于集中在某些部位。对于这一现象的一个合理的解释是交换较少发生在着丝粒的两旁。 同一染色体如果出现两个交叉,就交换所涉及的染色单体来讲,邻接的两个交换的关系有三种:包括两条染色单体的两线 双交换(图1a),包括三条染色单体的三线双交换(图1b、c),包括四条染色单体的四线双交换(图1d),它们分别带来1%、50%、100%基因重组;四分体类型分别属于包括两种类型的亲代二型(PD)、包括四种类型的四型(T)和包括两种类型的非亲代二型(NPD)。
一个位置上的一个交叉对于邻近位置上的交叉发生的影响称为干涉,如果一个交叉的发生减少另一交叉发生的概率则称为正干涉,如果增加后者发生的概率则称为负干涉。前一交叉影响邻近位置上另一任何形式的交叉的现象称为染色体干涉(交叉干涉);前一交叉所涉及的两条染色单体影响另一位置上相同的两条染色单体再次发生交换的现象称为染色单体干涉。 正交叉干涉的结果使实际发生的双交换频率低于假定两个交换的发生互不干涉所预期的频率。实际双交换频率与预期双交换频率之比称为并发系数。如果实际数值和理论值相同则并发系数是1,说明不存在交叉干涉。正交叉干涉的机制目前还不清楚。负交叉干涉现象曾在家蚕和λ噬菌体等中发现。一个位置上发生了一次交换以后,它的附近位置上更容易发生另一次交换。已经提出一些模型试图说明负交叉干涉的机制,但都还需要进一步验证。 如果不存在染色单体干涉,那么两线双交换、三线双交换和四线双交换频率之比应是1∶2∶1。染色单体干涉的结果使两线双交换减少,从而使子囊菌杂交子代中亲代二型子囊减少,或者使任何生物的杂交子代中重组型子代超过50%,这一现象曾在酵母菌和另一些真菌中发现。 发生的时间 在细胞减数分裂的过程中同源的两条染色体发生联会,然后各自分裂,形成包括四个染色单体的二价体 。在交叉位置上可以看到只有两条染色单体发生了交换(图1),这是说明交换发生在四线期的细胞学证据。遗传学证据来自四分体分析。如果交换发生在染色体分裂以前,那么通过减数分裂所产生的四分体将是非亲代二型(NDP),不发生交换的四分体将是亲代二型(PD),但是不可能出现四型(T)。只有交换发生在染色体已经分裂,即发生在四线期才有可能得到四型四分体。 关于交换发生的机制曾经有两种学说。一种学说称为样板选择,认为交换是染色单体复制过程中变换模版的结果。另一种学说称为断裂-重接,认为交换是复制完成后两个染色单体断裂并重新连接的结果。 影响因素 许多环境因素影响交换,包括温度、水分、营养、射线、化学药品等。很早便发现性别对于交换的影响,凡是性别为性染色体所控制的生物,异配性别个体中一般总是较少发生交换。1922年英国生理学家兼遗传学家J.B.S.霍尔丹提出了一条定律:凡是较少发生交换的个体必定是异配性别个体。这一定律称为霍尔丹定律。 交换和基因重组 如果基因在染色体上作线性排列,那么染色体交换应该带来基因重组。基因重组来源于同源染色体间的交换来说明。 由于交换发生在四线期,所以一次交换只导致半数染色单体上的基因发生重组。两线双交换并不带来重组,四线双交换带来全部染色单体的重组,三线双交换则带来50%重组,所以和一次交换一样,两次交换同样导致半数染色单体上的基因发生重组。三次、四次或更多次交换也只导致50%重组。如果两个基因不在同一染色体上,那么它们之间的组合必定是随机的,也就是说一个双基因杂合体的减数分裂产物中,亲本类型和重组类型各占50%。但是两个在同一染色体上的基因不论距离是多么远,重组值以50%为极限。所以重组值显然小于50%是基因连锁的证据,重组值是50%则并不是基因不连锁的证据。但是在两个相距较远的基因中间必然可以找到一些基因,它们和这两个基因的重组值将小于50%。整个连锁群就可以这样逐步建立。 自然群体中的交换和重组 每一种生物细胞中的染色体数是固定的,每一个细胞中全部染色体的交叉的平均数也是比较恒定的。交换的发生为基因所控制,交叉数的遗传控制是多基因的。交叉的位置也由基因所控制,这控制也是多基因的。交叉平均数大的群体的长期可塑性大,可是短期适应性较差;平均数小的群体则恰好相反。 交换除了受到基因的控制以外也和染色体结构有关。在果蝇中已经证明倒位杂合体中倒位能有效地抑制倒位区域内的交换。在许多果蝇群体中保持着这种染色体杂合状态,说明这种基因组合在特定生态条件下有利于生存。 染色体相互易位同样可以起到保持某种基因组合而减少重组的作用。在加州芍药和美洲蜚蠊中都曾发现某些自然群体中保持高度的易位杂合状态。一个极端的例子是一些拉马克月见草的染色体易位杂合体,这里易位所涉及的染色体是7对,在它们的减数分裂中期可以看到多达14个染色体形成的环,它们在中期的排列有90%属于交替式,并且交换都发生在染色体的两端。非但如此,它们的纯合体往往是致死的。这样一种平衡致死体系进一步保证杂合状态。并且实质上使所有的染色体成为一个连锁群,而很少可能通过交换而在连锁群间发生基因重组。 各种形式的交换和重组 体细胞交换于1936年首先由C.斯特恩在黑腹果蝇中发现。黄体(yellow body,y)和焦刚毛(singed, sn)这两个X连锁基因都呈杂合状态的雌性果蝇+y/sn+的表型是野生型。 可是在少数果蝇身上可以观察到孪生斑,即两小块邻接的组织:一块上显示突变型黄体性状,但上面的刚毛是正常的;旁边一块组织上的刚毛弯曲,但体色却是正常的。这一现象不能用基因突变,而只能用体细胞交换来解释(图2)。
交换一般是对等的,即两个同源染色体在对应的位置上发生交换,发生了交换的两个染色体都不增加或减少任何基因。如果同源染色体联会时不是准确地配对,那么交换发生在不对应的位置上,结果所得到的两个染色体中就有一个染色体缺少了一小部分,而另一染色体多了一小部分。这种交换称不等交换。通过不等交换所出现的这种多余的或重复的基因如果发生突变也不会影响细胞的生存,因为原有的基因可以执行正常的功能。这样,新的基因便可以通过多余基因的一再突变而形成。因此,不等交换被认为是进化过程中新基因的主要来源之一。 大肠杆菌和枯草杆菌都只有一个呈环状的染色体,即一个连锁群。在细菌染色体的重组过程中既不出现交叉,也没有联会线复合体,也不通过减数分裂而产生四分体。而且交换只发生在以单链形式进入受体的脱氧核糖核酸(DNA)片段中。尽管重组机制有所不同,但是真核生物和细菌染色体的重组都是以整段染色体的联会作为前提,这种重组方式称为同源重组或一般重组。 非同源重组是发生重组的两个部分中,它们只有极小一部分同源结构。它们在发生重组以前,不像减数分裂中的同源染色体那样整段染色体进行联会,而是联会仅发生在少到十几对核苷酸之间,而且重组的结果是一段 DNA插入到另一DNA分子中。 λ噬菌体的整合过程中并不进行复制,所以这种重组又称为保守性重组;转座子的整合过程包括它本身的复制,所以这种重组又称为复制式重组。 姊妹染色体单体互换 指来自两个亲本的染色体各自分裂成为两个姊妹染色单体后发生在姊妹染色单体之间的交换。早期的许多遗传学和细胞遗传学研究结果说明姊妹染色单体之间不发生交换。不过,至今还没有确切的证据足以证明姊妹染色单体互换在通过减数分裂发生的基因重组方面起着任何作用,它可能只是和DNA损伤有关。 连锁和交换的研究在实际应用中用于农业的育种和医学的产前诊断,在遗传学中是一个重要的研究课题。 |

说明:补充资料仅用于学习参考,请勿用于其它任何用途。
参考词条