1) evolution
[英][,i:və'lu:ʃn] [美]['ɛvə'luʃən]

进化
1.
Biological invasion and evolutionary biology of invasions;
生物入侵及入侵进化生物学
2.
Evolution of human T cell receptor variable gene segment families;
人类TCRαβ家族V基因的进化研究
3.
Human Hb s Failing to Interact on Human Hb and Its Evolutionary Significance;
几种鱼类血红蛋白不与人血红蛋白相互作用及其进化意义
2) evolvement
[英][i'vɔlvmənt] [美][ɪ'vɑlvmənt]
进化
1.
A Tentative Study on Optimization Theorem of Evolvement of Generic Self-organization;
一般自组织进化优化定理初探
2.
Beginning with the relationship between real man and nature,based on the analysis on modern strong doctrine of human cantered and why it becomes modern zoology calamity and environment crisis,the author points out that only by remolding the harmonious and evolvement relationship between man and nature,the harmonious,sustainable and healthy development of human society and nature can be actualized.
文章从现实的人与自然关系入手,在解读、分析现代“强式人类中心主义”及其何以构成现代生态灾难和环境危机的基础上,指出只有构建相对和谐、协调进化的人与自然关系,才能实现人类社会与自然的协调、持续、健康的发展。
3.
It is bring forward that the activity of human being have positive and negative actions to evolvements of earth.
就此本文予以讨论并提出人类活动对地球进化具有正反两方面作用。
3) evolutionary
[英][,i:və'lu:ʃənri] [美]['ɛvə'luʃə'nɛrɪ]
进化
1.
One of a random algorithm(WalkSAT),evolutionary algorithms and other heuristic algorithms are hot this year to study.
随着对SAT问题的深入研究,已经提出了很多高效的算法,其中随机算法(WalkSAT)、进化算法等启发式算法是今年来研究的热点。
2.
According to the problems of premature convergence and lower convergence speed in later evolutionary period of traditional GEP,an improved GEP algorithm is presented.
基因表达式编程(GEP)是基于遗传算法和遗传编程的具有更强数据处理和知识发现的进化算法。
3.
Evolutionary psychology is a new research orientation in western psychology, which makes uses of evolutionary theory to explore and study such issues as the origin and nature of psychology.
进化心理学是西方心理学研究中的一种新思潮,它运用进化理论探索人的心理的起源和本质。
4) Escalating evolution
递进进化
1.
A hybrid multi-objective genetic algorithm is proposed in this paper,which is based on operation-coded with an integration of escalating evolution and Pareto local search strategy.
通过将递进进化模式和非劣解局部搜索策略引入到算法结构,改善了算法对解空间的遍历性与算法的收敛性。
2.
The new algorithm takes a new elite duplication strategy and an innovative escalating evolutionary structure,which improved the convergence and efficiency of the algorithm and reduced its computational cost.
算法采用改进的精英复制策略,在实现精英保留的前提下降低了计算复杂性;通过递进进化模式增加群体多样性,改善了算法收敛性;通过群体进化过程中对非劣解集进行竞争型可变邻域启发式搜索,增强了算法局部搜索性能。
5) chemical evolution
化学进化
1.
Chemical reactions are the basis of chemical evolution.
无处不在的化学反应是化学进化的基础。
2.
Through chemical evolutions, proteins, lipids, triphosphate such as ATP and ribonucleic acid RNA are formed.
通过化学进化生成的蛋白质、脂质、三磷酸苷、核糖核酸等,在脂质组成的球膜内,形成具有能量转换、物质传递、并具有遗传性能的个体,是生命最初的形式。
3.
The origin of life is thought to start at the transit from chemical evolution into biological evolution.
生命起源开始于从化学进化进入生物进化。
6) Cultural evolution
文化进化
1.
Research on cultural evolutionary algorithms applied to Job shop problems;
Job shop调度文化进化算法研究
2.
Research on cultural evolutionary algorithms applied to Flow Shop problem with fuzzy delivery time
模糊交货期Flow Shop调度文化进化算法研究
3.
Based on analysis of the features of human being evolution,the thoughts that evolutionary computing simulates mechanism of biological evolution and(super-)group-evolutionary computing simulates mechanism of cultural evolution are proposed.
通过分析人类进化的特征,提出了进化计算模拟的是生物进化机制,(超)群体进化计算模拟的是文化进化机制的思想。
补充资料:进化
| 进化 evolution 19世纪后用于生物学,专指生物由简单到复杂、由低级到高级的变化发展。又称演化。evolution一词来自拉丁文evolutio,表示展开或把一个卷紧的卷松开的意思。 历史 达尔文以前生物变化思想的发展 关于万物互相转化和演变的自然观可以追溯到人类文明的早期。例如,中国《易经》中的阴阳、八卦说,把自然界还原为天、地、雷、风、水、火、山、泽八种基本现象,并试图用“阴阳”、“八卦”来解释物质世界复杂变化的规律。古希腊阿那克西曼德(约公元前6世纪)认为生命最初由海中软泥产生,原始的水生生物经过蜕变(类似昆虫幼虫的蜕皮)而变为陆地生物。 中世纪的西方,基督教圣经把世界万物描写成上帝的特殊创造物。这就是所谓特创论。与特创论相伴随的目的论则认为自然界的安排是有目的性的,“猫被创造出来是为了吃老鼠,老鼠被创造出来是为了给猫吃,而整个自然界创造出来是为了证明造物主的智慧”。从15世纪后半叶的文艺复兴到18世纪,是近代自然科学形成和发展的时期。这个时期在科学界占统治地位的观点是不变论。当时这种观点被I.牛顿和C.v林奈表达为科学的规律:地球由于所谓第一推动力而运转起来,以后就永远不变地运动下去on,生物种原来是这样,现在和将来也是这样。到了18世纪下半叶,I.康德的天体论首先在不变论自然观上打开了第一个缺口;随后,转变论的自然观就在自然科学各领域中逐渐形成。这个时期的一些生物学家,往往在两种思想观点中入门徬徨。例如林奈晚年在其《自然系统》一书中删去了物种不变的词句;法国生物学家G.-L.de布丰虽然把转变论带进了生物学 ,但他一生都在转变论和不变论之间徘徊。J.-B.de拉马克在1809年出版的《动物哲学》一书中详细阐述了他的生物转变论观点,并且始终没有动摇。 18世纪末~19世纪后期,大多数动植物学家都没有认真地研究生物进化,而且偏离了古希腊唯物主义传统,坠入唯心主义。“活力论”虽然承认生物种可以转变,但把进化原因归于非物质的内在力量,认为是生物的“内部的力量”即活力驱动着生物的进化,使之越来越复杂完善。但活力论缺乏实际的证据,是一种唯心的臆测。最有名的活力论者就是法国生物学家拉马克。19世纪后期出现的终极目的论或直生论,认为生物进化有一个既定的路线和方向而不论外界环境如何变化。 后人把拉马克对生物进化的看法称为拉马克学说或拉马克主义,其主要观点是:①物种是可变的,物种是由变异的个体组成的群体。②在自然界的生物中存在着由简单到复杂的一系列等级(阶梯),生物本身存在着一种内在的“意志力量”驱动着生物由低的等级向较高的等级发展变化。③生物对环境有巨大的适应能力;环境的变化会引起生物的变化,生物会由此改进其适应;环境的多样化是生物多样化的根本原因。④环境的改变会引起动物习性的改变,习性的改变会使某些器官经常使用而得到发展,另一些器官不使用而退化;在环境影响下所发生的定向变异,即后天获得的性状,能够遗传。如果环境朝一定的方向改变,由于器官的用进废退和获得性遗传,微小的变异逐渐积累,终于使生物发生了进化。拉马克学说中的内在意志带有唯心论色彩;后天获得性则多属于表型变异,现代遗传学已证明它是不能遗传的。 达尔文学说 1858年7月1日C.R.达尔文与A.R.华莱士在伦敦林奈学会上宣读了关于物种起源的论文。后人称他们的自然选择学说为达尔文-华莱士学说。达尔文在1859年出版的《物种起源》一书中系统地阐述了他的进化学说。其核心自然选择原理的大意如下:生物都有繁殖过剩的倾向,而生存空间和食物是有限的,所以生物必须“为生存而斗争”。在同一种群中的个体存在着变异,那些具有能适应环境的有利变异的个体将存活下来,并繁殖后代,不具有有利变异的个体就被淘汰。如果自然条件的变化是有方向的,则在历史过程中,经过长期的自然选择,微小的变异就得到积累而成为显著的变异。由此可能导致亚种和新种的形成。 达尔文的进化理论,从生物与环境相互作用的观点出发,认为生物的变异、遗传和自然选择作用能导致生物的适应性改变。它由于有充分的科学事实作根据,所以能经受住时间的考验,百余年来在学术界产生了深远的影响。但达尔文的进化理论还存在着若干明显的弱点:①他的自然选择原理是建立在当时流行的“融合遗传”假说之上的。按照融合遗传的概念 ,父、母亲体的遗传物质可以像血液那样发生融合;这样任何新产生的变异经过若干世代的融合就会消失,变异又怎能积累、自然选择又怎能发挥作用呢②达尔文过分强调了生物进化的渐变性;他深信“自然界无跳跃”,用“中间类型绝灭”和“化石记录不全”来解释古生物资料所显示的跳跃性进化。他的这种观点近年正越来越受到间断平衡论者和新灾变论者的猛烈批评。 达尔文以后进化论的发展 1865年奥地利植物学家G.J.孟德尔从豌豆的杂交实验中得出了颗粒遗传的正确结论。他证明遗传物质不融合,在繁殖传代的过程中,可以发生分离和重新组合。20世纪初遗传学建立,T.H.摩尔根等人进而建立了染色体遗传学说,全面揭示了遗传的基本规律。这本应弥补达尔文学说的缺陷,有助于进化论的发展;但当时大多数遗传学家(包括摩尔根在内),都反对达尔文的自然选择学说。人们对达尔文进化论的信仰,发生了严重的危机。 ①新拉马克主义与新达尔文主义。在19世纪末到20世纪初这个时期出现过一些新的进化学说。荷兰植物学家H.德·弗里斯在20世纪初根据月见草属的变异情况提出“物种通过突变而产生”的突变论,而反对渐变论。这个理论得到当时许多遗传学家的支持。某些拉马克学说的追随者们虽然抛弃了拉马克的内在意志概念,但仍强调后天获得性遗传,并认为这是进化的主要因素。50年代在苏联由T.D.李森科所标榜的米丘林学说,强调生物在环境的直接影响下能够定向变异、获得性能够遗传。所有这些观点被称为新拉马克主义。A.F.L.魏斯曼在1883年用实验来证明获得性遗传的错误,强调自然选择是推动生物进化的动力,他的看法被后人称为新达尔文主义。 ②现代综合进化学说。20世纪20~30年代首先由R.A.费希尔、S.赖特和J.B.S.霍尔丹等人将生物统计学与孟德尔的颗粒遗传理论相结合,重新解释了达尔文的自然选择学说,形成了群体遗传学。以后C.C.切特韦里科夫、T.多布然斯基、J.赫胥黎、E.迈尔、F.J.阿亚拉、G.L.斯特宾斯、G.G.辛普森和J.W.瓦伦丁等人又根据染色体遗传学说、群体遗传学、物种的概念以及古生物学和分子生物学的许多学科知识,发展了达尔文学说,建立了现代综合进化论。现代综合进化论彻底否定获得性状的遗传,强调进化的渐进性,认为进化是群体而不是个体的现象,并重新肯定了自然选择的压倒一切的重要性,继承和发展了达尔文进化学说。 ③中性学说和间断平衡论。1968年,日本学者木村资生根据分子生物学的材料提出了分子进化的中性学说(简称中性学说)。认为在分子水平上,大多数进化改变和物种内的大多数变异,不是由自然选择引起的,而是通过那些选择上中性或近乎中性的突变等位基因的随机漂变引起的,反对现代综合进化论的自然选择万能论观点(见分子进化的中性学说)。 1972年N.埃尔德雷奇和S.J.古尔德共同提出“间断平衡”的进化模式来解释古生物进化中的明显的不连续性和跳跃性,认为基于自然选择作用的种以下的渐进进化模式,即线系渐变模式,不能解释种以上的分类单元的起源,反对现代达尔文主义的唯渐进进化观点。争论仍在继续中(见间断平衡论)。 小进化与大进化 美国遗传学家R.B.戈德施米特认为,通常的自然选择,只能在物种的范围内,作用于基因而产生小的进化改变,即小进化;而由一个种变为另一个种的进化步骤则需要另一种进化方式,即大进化。他认为大进化就是通过他所假设的系统突变(涉及整个染色体组的遗传突变)而实现的。这样就可以一下子产生出一个新种甚至一个新属或新科。美国古生物学家辛普森同意把进化的研究分成两大领域:研究种以下的进化改变的小进化和研究种以上层次的进化的大进化,但并不同意戈氏的观点,他并不认为小进化与大进化是各自不同的或彼此无关的进化方式。 小进化研究种以下的进化改变,包括:①小进化的因素和机制,研究遗传突变、自然选择、随机现象(如遗传漂变)等因素如何引起群体的遗传组成的改变等。②种形成,研究新种的形成方式和过程,研究小进化因素如何导致同种的群体之间的隔离的形成和发展,研究种内分化和由亚种、半种到完全的种的发展过程等。 大进化研究种以上的分类单元在地质时间尺度上的进化改变,其对象主要是化石,最小研究单位是种。主要研究内容包括:①种及种以上分类单元的起源和大进化的因素。②进化型式,在时间向度上进化的线系的变化和形态。③进化速度,形态改变的速度和分类单元的产生或绝灭速度,种的寿命等。④进化的方向和趋势。⑤绝灭的规律、原因及其与进化趋势、速度的关系等。 小进化与大进化在物种这一层次上相互衔接,事实上小进化与大进化都研究物种形成。关于小进化与大进化的关系问题,近年学术界展开了激烈的争论。间断平衡学派认为不能以小进化的机制来解释大进化的事实;而现代综合进化论则认为小进化是大进化的基础,小进化的机制在一定程度上是可以说明大进化的现象的。 进化型式 从进化的观点看,同时生存的不同生物种在时间的向度上可以回溯到一个共同祖先。因此,按照祖裔关系可以将现时生存的和曾经生存过的生物相互联结起来,这种表示祖裔关系的生物进化系统称为种系发生。生物的种系发生可以形象地表示为一棵树:如果从树根到树顶代表时间向度,主干代表共同祖先,大小分枝代表相互关联的进化线系,这就构成所谓种系发生树或进化树。所谓进化型式就是进化在时间与空间上的特征,也就是种系发生的特征,具体表现在进化树的形态上:枝干的延续和分枝方式、树干的倾斜方向和在空间上的配置、树干的中断等等,它代表着种系发生中线系进化、种形成、绝灭等方面的特征。 在谱系进化中有两种性质不同的进化改变。一是形态结构及其功能由简单、相对不完善到复杂和相对完善的前进性(进步的)改变,称之为前进进化;前进进化的结果是造成生物的等级从低级到高级。另一种进化改变是线系分枝,叫做分枝进化;分枝进化的结果是产生新的分类单元和生物歧异度的增长。广义的前进进化包括除分枝进化以外的各种进化改变,既包括前进的(进步的)进化改变,也包括非前进的甚至退化的改变。既无前进进化,又无分枝进化的情况称为停滞进化,如活化石之类的情况。 适应辐射 在相对较短的地质时间内,从一个线系分枝出许多歧异的分类单元,叫做辐射。由于辐射分枝通常是向不同的方向适应进化的,所以又称为适应辐射。 趋同进化与平行进化 不同的线系各自独立地进化出相似的特征叫做趋同进化,简称趋同。形态结构的进化趋同往往是由于功能的相似,而功能的相似又往往是由于适应于相似的环境。 平行进化(简称平行)是指两个或多个有共同祖先的线系,在其祖先遗传的基础上分别独立进化出相似的特征。通常平行与趋同不易区分,一般说来平行进化既涉及同功又涉及同源;假若后裔间的相似程度大于各自的祖先之间的相似程度则可称为趋同,若后裔之间的相似程度与其祖先之间的相似程度差不多则可称为平行(图1)。
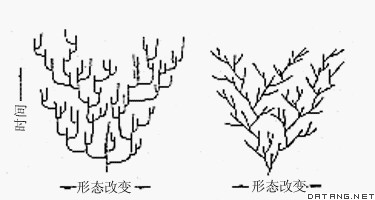
大进化的模式 ①渐变模式。认为形态进化速度多少是恒定的、匀速的,形态进化是逐渐的;形态改变主要是线系进化造成的,大多发生在种的生存期间,与种形成无关,种形成(线系分枝)本身只是增加新的进化方向。②间断模式。认为形态进化速度是不恒定、非匀速的,快速的“跳跃”与长期的停顿相交替,即在种形成期间进化加速,种形成后保持相对的稳定。进化的形态改变与种形成相关联,即大部分形态改变是发生在相对较短的种形成时期,在可能长达数百万年的种生存期内不会发生显著的形态改变(图2)。两种模式各有一些有利于自己的证据,但这些证据本身还需进一步研究才能证实。
进化速率 单位时间内生物进化改变量。衡量进化速率必须确定两个尺度:时间尺度和进化改变量的尺度。时间尺度有两种,即绝对地质时间和相对地质时间,一般应使用绝对时间尺度;只在不得已时,才使用相对时间尺度。 进化趋势 从长的时间尺度来看生物进化呈某种方向性,但这并非说自然界存在着既定的进化轨道,这里说的方向性是统计学的趋向。因此,进化趋势是任何因素(包括随机的非随机的因素)所造成的进化的统计学趋势。 小进化趋势 小进化趋势是指在生物种的自然群体之内,自然选择作用所造成的表型进化改变的趋向性。这种进化趋势通常是适应局部环境的结果,进化改变发生在线系之内,所以也可称之为线系进化趋势,表现为线系在时间-进化改变坐标系内的倾斜。 大进化趋势 大进化趋势是指在种系发生中线系分枝的主体的趋向性。大进化趋势发生在种以上的分类单元的谱系进化中,所以又可以称为谱系进化趋势,表现为谱系中的偏斜。为解释大进化趋势发生的原因,不少学者提出过种种假说,但由于缺乏证据,能够成立或有说服力的还很少见。 |

说明:补充资料仅用于学习参考,请勿用于其它任何用途。
参考词条